

Clusterin (Apolipoprotein J): An Extracellular Chaperone Linking Proteostasis, Biomarker Science, and Precision Medicine
1. Introduction
Clusterin (CLU), also known as apolipoprotein J, is a secreted glycoprotein known primarily to function as an ATP-independent extracellular molecular chaperone (Du et al., 2025). It is found in high abundance in bodily fluids such as plasma and cerebrospinal fluid (CSF), making it clinically detectable and biologically significant (Yuste-Checa et al., 2025). Unlike most intracellular chaperones, clusterin is closely associated with the maintenance of extracellular proteostasis and processing of damaged or improperly folded proteins outside the cell (Carini et al., 2025). As a result, it has been implicated in several diseases, including cancer, neurodegenerative disease, kidney disease, metabolic disorders, and tissue injury (Sultana and Novotny, 2024). This protein is no longer viewed solely as a stress-response protein but also as a disease biomarker. Recent research suggests that clusterin may act as a disease-related biomarker, a tissue-protective factor, and, in certain circumstances, a therapeutic target (Kalvaityte et al., 2022). In addition, its biology is more complex, as it exists in multiple forms, locations, and glycosylation states that can affect its function (Janiszewska et al., 2022). This review discusses clusterin from a protein-centered perspective, with emphasis on its structural characteristics, biosynthesis, biological functions, clinical relevance, direct detection methods, and translational potential.2. Molecular structure and architectural features of clusterin
Mature clusterin is a heavily glycosylated heterodimer composed of α and β chains linked by disulfide bonds. This structural organization contributes to the protein’s stable, soluble, and extracellular chaperone function. Glycosylation is also important because it affects protein movement, molecular interactions, and disease-related variability (Janiszewska et al., 2022). Clusterin is therefore understood not only by its amino acid sequence, but also by its glycan composition (Liang et al., 2015). As shown in Figure 1, the wild-type clusterin model highlights its α- and β-chains and its glycan-rich structure, which help explain its stability and broad extracellular chaperone activity. Recent evidence further showed that human clusterin has a discontinuous three-domain architecture with flexible hydrophobic tails that are important for client binding and chaperone action (Yuste-Checa et al., 2025). This advance moved the field beyond the past descriptive models and provided a clear explanation of how clusterin interacts with amyloidogenic and non-native proteins (Du et al., 2025). These structural findings also help explain why clusterin can interact with receptors, lipids, and aggregation-prone proteins without functioning as a classical enzyme with a single, narrow catalytic function (Carini et al., 2025).
Figure 1. The structural model of wild-type human clusterin.
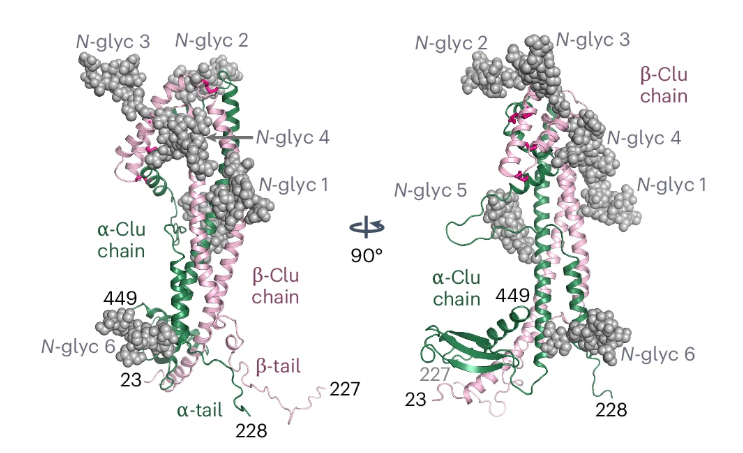
(Yuste-Checa et al., 2025)
3. Biogenesis and cellular processes of clusterin
Clusterin is synthesized as a single precursor that enters the secretory pathway, undergoes signal peptide removal, acquires multiple N-glycans and is later cleaved into α and β chains that remain linked by disulfide bonds. It is produced via the classical endoplasmic reticulum and Golgi apparatus secretory route, which corresponds to its' predominant extracellular role (Yuste-Checa et al., 2025). However, its biology is not limited to the secreted form, as high levels of cellular stress can alter localization and generate intracellular pools with distinct functions (Satapathy et al., 2023). This is important because secreted clusterin is associated with extracellular quality control, whereas nuclear forms are linked to altered stress handling, protein degradation, and cell fate determination (Du et al., 2025). These findings help explain why clusterin could be protective in some diseases but harmful in others (Sultana and Novotny, 2024). Furthermore, recent evidence demonstrates that endoplasmic reticulum stress can cause the release of clusterin into the cytoplasm, where it can help direct misfolded proteins towards degradation pathways rather than allowing toxic aggregates (Satapathy et al., 2023). These findings extend clusterin’s role from being only a secreted holdase-like chaperone to also a more flexible proteostasis regulator during stress (Carini et al., 2025).
4. Physiological roles and functional properties of clusterin
Clusterin’s well-known biological function is to bind unfolded, damaged, or aggregation-prone extracellular proteins, thereby keeping them soluble and less toxic (Yuste-Checa et al., 2025). This is particularly important for tissues under chronic mechanical stress, as aggregated proteins can damage receptor membranes and surrounding cells (Carini et al., 2025). Clusterin is also linked to lipid transport and lipoprotein-like behavior, expanding its roles beyond chaperoning (Du et al., 2025). Additionally, clusterin may affect complement-related injury, resistance to apoptosis, and tissue repair, although the exact effect depends on the disease setting and the protein form involved (Sultana and Novotny, 2024). In neurology, clusterin has gained strong attention due to its interactions with proteins and processes implicated in Alzheimer’s and Parkinson’s disease (Yu et al., 2025). Secretory clusterin decreased dopaminergic neuron apoptosis and retained protective cellular functions in experimental models of Parkinson’s disease, demonstrating a neuroprotective role (Zhu et al., 2024). In metabolic tissues, circulating clusterin has also been associated with adipose tissue insulin resistance, suggesting that clusterin biology extends to systemic metabolic dysfunction (Wang et al., 2023). These findings show that clusterin is not a niche protein; it connects extracellular stress control to broader organ-level disease biology (Mao et al., 2025).
5. Clusterin in disease and biomarker research
Clusterin has become particularly attractive in biomarker research because it is measurable in readily accessible biofluids and is altered in several chronic diseases (Kalvaityte et al., 2022). In kidney disease, urinary clusterin has shown strong value as a marker of epithelial stress and senescence, and recent human studies have linked higher urinary clusterin levels to poorer renal outcomes (Baird et al., 2025). Recent studies further supported urinary clusterin as a clinically meaningful kidney biomarker by linking it to disease progression and treatment-response biology in diabetic chronic kidney disease (Ju et al., 2026). This makes clusterin more compelling than many theoretical biomarkers, as it is already moving toward translational utility (Mao et al., 2025).
Recent clinical evidence showed that serum clusterin levels are elevated in women with polycystic ovary syndrome, supporting its value as a measurable biomarker in reproductive and metabolic disorders. As shown in Figure 2, ROC analysis demonstrated that clusterin had useful diagnostic performance in distinguishing polycystic ovary syndrome (PCOS), alongside established markers such as AMH and the LH/FSH ratio. These findings strengthen the view that clusterin is not only biologically relevant in disease but also has potential clinical value as a serum biomarker (Kurt et al., 2026). Beyond kidney and reproductive disease, clusterin has also shown biomarker relevance in oncology, where CLU expression has been linked to prognosis and tumor-related biology (Fu et al., 2023). At the same time, the literature repeatedly warns that clusterin can be protective in some tissues and pro-survival in some tumors, so interpretation must remain disease-specific (Sultana and Novotny, 2024).
Figure 2. Diagnostic performance of serum clusterin in women with polycystic ovary syndrome.
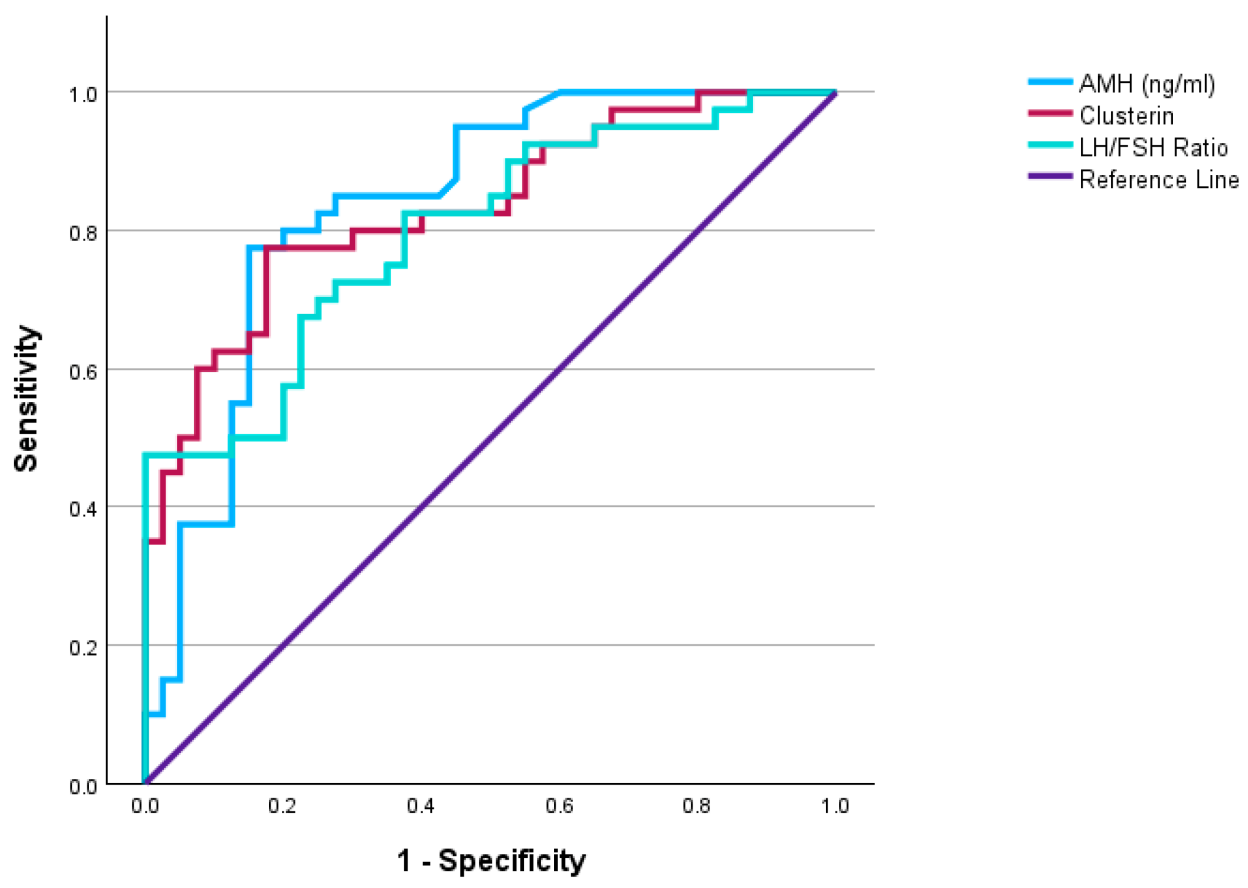
(Kurt et al., 2026)
6. Therapeutic potential of clusterin
Clusterin is an interesting therapeutic target because altering its levels may modulate proteotoxic stress, tissue injury, and disease progression (Carini et al., 2025). In neurodegeneration, one promising direction is to enhance protective secreted clusterin rather than to suppress it (Yu et al., 2025). This idea gained strong support when a small-molecule secreted clusterin enhancer increased brain clusterin and improved memory-related outcomes in Alzheimer’s disease mouse models (Cohn et al., 2025). Such findings show that clusterin-targeted therapy does not always mean inhibition. In some diseases, it may mean restoring a protective extracellular defense system (Zhu et al., 2024). The opposite may be true in some cancers, where clusterin can support cell survival, treatment resistance, or tumor adaptation (Sultana and Novotny, 2024). This explains why clusterin is best understood as a context-dependent therapeutic target rather than a universally good or bad protein (Du et al., 2025). Future clusterin therapies will likely depend on isoform awareness, tissue targeting, and careful patient selection (Fu et al., 2023).
7. Analytical approaches for clusterin detection
Direct measurement of clusterin in biofluids is most often performed using immunoassays, especially ELISA-based methods. This has been used successfully in tears, urine, serum, plasma, and cerebrospinal fluid, depending on the clinical question (Yu et al., 2018). In kidney disease, ELISA quantified urinary clusterin in large patient cohorts and linked it with outcome prediction (Baird et al., 2025). In reproductive medicine, serum clusterin is also directly measured and evaluated through ROC analysis in women with PCOS (Kurt et al., 2026). When tissue localization is needed, Western blotting, immunohistochemistry, and immunofluorescence are the most informative direct protein methods because they show both abundance and anatomical distribution. For example, recent work on keloids validated clusterin changes using Western blot and IHC in patient-derived material, demonstrating how tissue-level detection can complement serum studies (Niu et al., 2025).
In kidney studies, paired-biopsy immunofluorescence has also been used to link urinary clusterin to senescent renal epithelial cells within the tissue itself (Baird et al., 2025). Mass spectrometry is important when the goal is to separate clusterin isoforms or glycoforms rather than only to quantify total protein. This is essential because clusterin is heavily glycosylated, and disease-associated differences may be hidden if only total immunoreactivity is measured (Liang et al., 2015). Recent discussions of clusterin glycosylation have therefore emphasized glycoproteomics as a high-value approach for biomarker refinement (Janiszewska et al., 2022). Emerging technologies such as SERS-based immunoassays are also promising because they may allow sensitive detection with small sample volumes. A recent study reported the detection of clusterin down to about 3 ng/mL on an aluminum foil substrate (Mergenbayeva et al., 2025).
8. Conclusions
Clusterin is a multifunctional glycoprotein with important roles in extracellular proteostasis and protection against cellular stress. Its structure, biosynthesis, and multiple protein forms explain its broad biological activity. Current evidence shows that clusterin is relevant in neurodegenerative, renal, metabolic, reproductive, and cancer-related conditions. Its detectability in body fluids also supports its value as a promising biomarker. Overall, clusterin is both biologically important and clinically relevant for future diagnostic and therapeutic research.
References
Baird, D.P., Reck, M., Campbell, R., Docherty, M.-H., Janas, P.P., Mason, T., Mortuza, Z., Vermeren, M., Nam, A., Yang, W., Schurman, N., Williams, C., Traynor, J.P., Mark, P.B., Mylonas, K.J., Hughes, J., Denby, L., Conway, B.R. and Ferenbach, D.A. (2025). Urinary Clusterin is a Biomarker of Renal Epithelial Senescence and Predicts Human Kidney Disease Progression. Kidney International Reports, 10(7), pp. 2344–2356. https://doi.org/10.1016/j.ekir.2025.04.035
Carini, G., Mohammed, S., Filippini, A., Ramazzina, I. and Russo, I. (2025). The Emerging Role of the Molecular Chaperone Clusterin in Parkinson’s Disease. International Journal of Molecular Sciences, 26(13), 6351. https://doi.org/10.3390/ijms26136351
Cohn, W., Campagna, J., Wi, D., Lee, J.T., Beniwal, S., Elezi, G., Zhu, C., Jagodzinska, B., Whitelegge, J., Damoiseaux, R. and John, V. (2025). Discovery of a small molecule secreted clusterin enhancer that improves memory in Alzheimer’s disease mice. npj Drug Discovery, 2, 7. https://doi.org/10.1038/s44386-025-00009-2
Du, X., Chen, Z. and Shui, W. (2025). Clusterin: structure, function and roles in disease. International Journal of Medical Sciences, 22(4), pp. 887–896. https://doi.org/10.7150/ijms.107159
Fu, Y., Du, Q., Cui, T., Lu, Y. and Niu, G. (2023). A pan-cancer analysis reveals role of clusterin (CLU) in carcinogenesis and prognosis of human tumors. Frontiers in Genetics, 13, 1056184. https://doi.org/10.3389/fgene.2022.1056184
Janiszewska, E., Kmieciak, M., Waszak, P.M., Polańska, J., Mazurek, U. and Świętochowska, E. (2022). The Influence of Clusterin Glycosylation Variability on Its Function in Lipid Transportation and Aβ Toxicity. BioMed Research International, 2022, 7657876. https://doi.org/10.1155/2022/7657876
Ju, W., Nair, V., Vart, P., et al. (2026). Urinary clusterin as a biomarker of human kidney disease progression and response to the endothelin receptor antagonist atrasentan: An exploratory analysis from the SONAR trial. Nature Communications, 17, Article 68973. https://doi.org/10.1038/s41467-026-68973-5
Kalvaityte, U., Matta, C., Bernotiene, E., Pushparaj, P.N., Kiapour, A.M. and Mobasheri, A. (2022). Exploring the translational potential of clusterin as a biomarker of early osteoarthritis. Journal of Orthopaedic Translation, 32, pp. 77–84. https://doi.org/10.1016/j.jot.2021.10.001
Kurt, D.S., Ağaoğlu, R.T., Kıncı, M.F., Sırma, T., Kurt, A., Pay, R.E., Gültekin, İ.B., Keskin, H.L. and Aksakal, S.E. (2026). Diagnostic Value and Metabolic Association of Serum Clusterin in Women with Polycystic Ovary Syndrome. Diagnostics, 16(1), 167. https://doi.org/10.3390/diagnostics16010167
Liang, H.-C., Russell, C., Mitra, V., Chung, R., Hye, A., Bazenet, C., Lovestone, S. and Soininen, H. (2015). Glycosylation of Human Plasma Clusterin Yields a Novel Candidate Biomarker of Alzheimer’s Disease. Journal of Proteome Research, 14(12), pp. 5063–5076. https://doi.org/10.1021/acs.jproteome.5b00892
Mao, L., Yin, R., Yang, L. and Zhao, D. (2025). Elucidating the function of clusterin in the progression of diabetic kidney disease. Frontiers in Pharmacology, 16, 1573654. https://doi.org/10.3389/fphar.2025.1573654
Mergenbayeva, S., Terzapulo, X. and Bukasov, R. (2025). Detection of Biomarker Clusterin in SERS Immunoassays on Al Foil After Substrate Selection and Assay Optimization with Fluorescently Labeled Antibodies. Molecules, 30(19), 3974. https://doi.org/10.3390/molecules30193974
Niu, Y., Zhang, X., Chen, Y. and Yao, X. (2025). Secreted clusterin inhibits keloid formation by promoting fibroblast apoptosis. Molecular and Cellular Probes, 83, 102047. https://doi.org/10.1016/j.mcp.2025.102047
Satapathy, S., Walker, H., Brown, J., Gambin, Y. and Wilson, M.R. (2023). The N-end rule pathway regulates ER stress-induced clusterin release to the cytosol where it directs misfolded proteins for degradation. Cell Reports, 42(9), 113059. https://doi.org/10.1016/j.celrep.2023.113059
Sultana, P. and Novotny, J. (2024). Clusterin: a double-edged sword in cancer and neurological disorders. EXCLI Journal, 23, pp. 912–936. https://doi.org/10.17179/excli2024-7369
Wang, Y., Yu, H., Ma, X., Wang, Y., Liu, W., Zhang, H., Chen, W., Yu, S., Bao, Y. and Yang, Y. (2023). Clusterin is closely associated with adipose tissue insulin resistance. Diabetes/Metabolism Research and Reviews, 39(7), e3688. https://doi.org/10.1002/dmrr.3688
Yu, V., Bhattacharya, D., Webster, A., Bauskar, A., Flowers, C., Heur, M., Chintala, S.K., Itakura, T., Wilson, M.R., Barr, J.T., Jeong, S., Wang, M. and Fini, M.E. (2018).Clusterin from human clinical tear samples: Positive correlation between tear concentration and Schirmer strip test results. The Ocular Surface, 16(4), pp. 478–486. https://doi.org/10.1016/j.jtos.2018.08.001
Yu, Y., Wang, C., Wang, B., Wang, X., Zhao, Q., Yan, Y. and Liu, X. (2025). Clusterin Regulates the Mechanisms of Neuroinflammation and Neuronal Circuit Impairment in Alzheimer’s Disease. International Journal of Molecular Sciences, 26(15), 7271. https://doi.org/10.3390/ijms26157271
Yuste-Checa, P., Carvajal, A.I., Mi, C., Paatz, S., Hartl, F.U. and Bracher, A. (2025). Structural analyses define the molecular basis of clusterin chaperone function. Nature Structural & Molecular Biology, 32, pp. 2035–2045. https://doi.org/10.1038/s41594-025-01631-4
Zhu, D., Zhang, S., Wang, X., Xiao, C., Cui, G. and Yang, X. (2024). Secretory Clusterin Inhibits Dopamine Neuron Apoptosis in MPTP Mice by Preserving Autophagy Activity. Neuroscience, 540, pp. 38–47. https://doi.org/10.1016/j.neuroscience.2024.01.010








